Прекурсори синтезата на L-синџирот на антитела. Ekstrapeptidy и нивната функција
Претходник L-синџирот на може да се открие во систем за клеточно-слободен содржи полирибозоми од MOPC 21 клетки, додека во системи кои содржат микрозоми (Milstein e. a., 1972) или жаба ооцити (Фауст e. a., 1974), откриени само зрели L-синџир. Така, конверзија на прекурсор во L-синџир, се претпоставува дека е во функција на ендоплазматичниот ретикулум. Неодамна, синтезата на прекурзорот L-синџирот беше во можност да се идентификуваат и цели клетки (Schmeckpeper д. A., 1975).
Во моментов, истрага структура на неколку прекурзори. Компаративна големини и делумна секвенца на амино киселини во NH2-терминал ekstrapeptidah претходник лесни ланци на имуноглобулини (Burstein e. А., 1976- Burstein, Шехтер, 1976) се претставени подолу.
Покажано е дека тие содржат 19 или 20 амино киселини повеќе од зрели L-tsepi- NH2-терминал амино киселина е метионин. Карактеристика на сите NH2-терминал ekstrapeitidov на висока содржина на хидрофобни амино киселини (50-70%) што го прави нивната изречена хидрофобен карактер. Така, во ekstrapeptidah на L-синџирот на SIDS SIDS 63 и 321 има две леуцин тројки во MOPC 41 има kvadriplet, и MOPC 104E-5 тесно распоредени остатоци се од оваа амино киселина. Вреди да се спомне дека податоците кои неодамна беа добиени кои им овозможуваат да се претпостави постоење на дополнителни пептид, а исто така и на COOH koitse L-синџири (Шехтер д. A., 1975), но ова е неизвесна.
Познато е дека на L-синџир SIDS SIDS, а 63 321 се на истиот подгрупа, и нивните V-регионот се разликуваат од само 8 амино киселини од ekstrapeptidy 111. Според тоа, овие кола се идентични, или барем многу слични. Во контраст, L-DITS MOPC 41 кои припаѓаат на друга подгрупа kappa синџири се разликуваат во нивните V-домени од L-tseiey MOPC 63 и MOPC 321 на 46 и 48%, соодветно, и нивните ekstrapeptidy се разликуваат од ekstrapeptidov MOPC 63 и MOPC 321 40 %. Значаен разлики (не помалку од 7 амино киселина од 20), исто така, се забележани помеѓу ekstraneptidami капа и ламбда синџири. Во исто време, тоа може да изгледа да се предвиди дека Л-ekstrapeptidy tseney Nors 2020 разликуваат од L-синџири MOPC 104E само две амино киселини од V-региони ќе бидат идентични.

Во присуство на метионин само на крајот на L-синџирот на прекурсор-NH2 тоа покажува дека овие молекули се директен производ mRNA превод, почнувајќи од нејзиниот крај-5`. Ова е потврдено од страна на фактот дека на mRNA лишен 5`-koitsa, во систем ќелија бесплатни не се емитува (Matthyssens д. А., 1976- Shimotohno д. A., 1977). Сите овие земени заедно укажува на тоа дека NH2-завршен дел е ekstrapeptid V-регионот и дека, следствено, V-ген подолго од претходно предвидено.
функции ekstrapeptidov сè уште нејасна. Дали тие се состои во обезбедување синтетизира синџири интеракција со мембрани на ендоплазматичниот ретикулум, а можеби и на површината на клетката. Ако тоа е точно, тогаш ekstrapeptidy можат да служат дополнителен механизам за признавање. Од друга страна, пак, според податоците Milstein et al (Milstein д. A., 1972) во микрозомални мембрана содржи ензим пресекување ekstrapeptidy брзо. Ова укажува на тоа дека тие играат улога само во почетните фази на синџирот на трансфер вектор во микрозомални везикули.
Очигледно, таа треба да очекуваме NH-откривање терминал ekstrapeptidov и H-синџири. Некои показател за ова е откривањето на необични пептиди во H-синџири на IgA ламбда-тип (Barstad д. A., 1974). и IgG3 (Dammaco e. a., 1972).
исклучително интересни Таа, исто така ќе провери дали има ekstrapeptidy не само mielompyh, но во нормални имуноглобулини и антитела, и дали тие се разликуваат во различни антитела со специфичност. Досега испитани само еден миелома протеини од активноста на антитела - MOPC 104E.
Сите од горенаведените укажува на тоа дека mRNA превод ин витро системи мобилен бесплатни доведе до откривање на претходно непознати фаза на синтеза на имуноглобулини и постави голем број на нови прашања од страна на истражувачите.
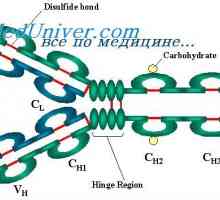 Имуноглобулин лесни ланци. организација на имуноглобулини
Имуноглобулин лесни ланци. организација на имуноглобулини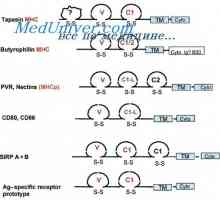 Контрола на образование имуноглобулин лесни ланци. Гените лесни ланци антитело
Контрола на образование имуноглобулин лесни ланци. Гените лесни ланци антитело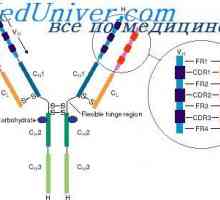 Локализација на гени лесни ланци антитело. Причините за променливоста на лесни ланци на
Локализација на гени лесни ланци антитело. Причините за променливоста на лесни ланци на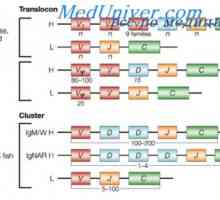 Translocon имуноглобулини. Комбинирање V- и гените на антитела
Translocon имуноглобулини. Комбинирање V- и гените на антитела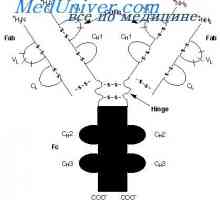 Разликите варијабилни региони на антитела. Променлив подгрупа имуноглобулин
Разликите варијабилни региони на антитела. Променлив подгрупа имуноглобулин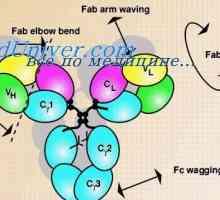 На варијабилни региони на антитела тешки ланци. Варијабилен регион гените на тешки ланци
На варијабилни региони на антитела тешки ланци. Варијабилен регион гените на тешки ланци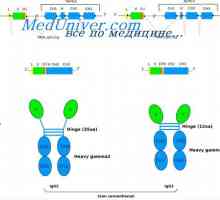 Poliribosomny комплекс синтеза на антитела. RNA кои се вклучени во синтеза на антитела
Poliribosomny комплекс синтеза на антитела. RNA кои се вклучени во синтеза на антитела Формирање на имуноглобулин синџири. Вишокот антитело синтеза l-синџири
Формирање на имуноглобулин синџири. Вишокот антитело синтеза l-синџири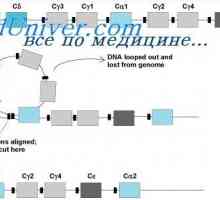 Методи за изолација на полирибозоми. Димензии полирибозоми синтетизираат антитела
Методи за изолација на полирибозоми. Димензии полирибозоми синтетизираат антитела Синтеза на имуноглобулин со тежок и лесни ланци. Единствен синтеза на тешки и лесни ланци антитело
Синтеза на имуноглобулин со тежок и лесни ланци. Единствен синтеза на тешки и лесни ланци антитело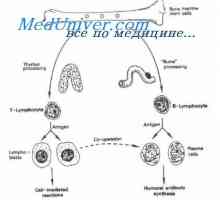 MRNA е вклучен во синтезата на антитело. Методи на проучување на mRNA
MRNA е вклучен во синтезата на антитело. Методи на проучување на mRNA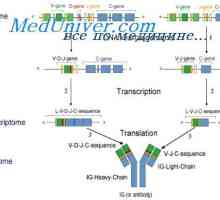 Properties антитела mRNA. MRNA структура на имуноглобулини
Properties антитела mRNA. MRNA структура на имуноглобулини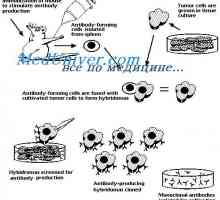 Превод на mRNA. Карактеристики на mRNA превод под синтеза на антителото
Превод на mRNA. Карактеристики на mRNA превод под синтеза на антителото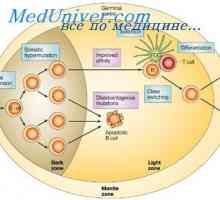 Плазма глувци тумори. Полирибозоми улога во биосинтезата на имуноглобулини
Плазма глувци тумори. Полирибозоми улога во биосинтезата на имуноглобулини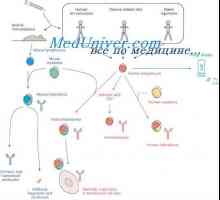 Хибридизација на mRNA и ДНК антитела. Локализација V- и гени во геномот на имуноглобулини
Хибридизација на mRNA и ДНК антитела. Локализација V- и гени во геномот на имуноглобулини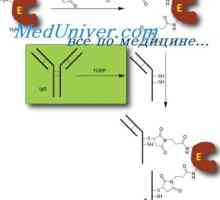 Составување на антитела. комплементирање на имуноглобулини
Составување на антитела. комплементирање на имуноглобулини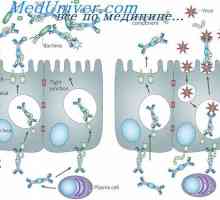 На секреција на имуноглобулини. Фази секреција антитело
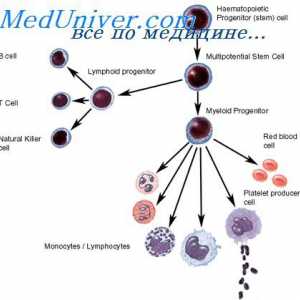
На секреција на имуноглобулини. Фази секреција антитело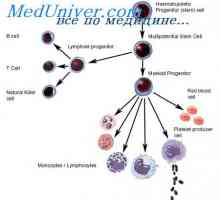 Образование лимфоцити прекурзори. Лезии на матични клетки
Образование лимфоцити прекурзори. Лезии на матични клетки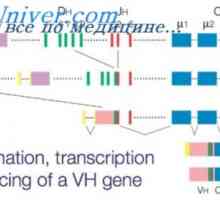 Ефектот на фенотип на висока синџирот на антитела. Ограничување V-гени
Ефектот на фенотип на висока синџирот на антитела. Ограничување V-гени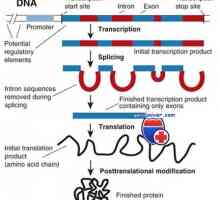 Транскрипција. Форми и видови на РНК клетки
Транскрипција. Форми и видови на РНК клетки Амино киселини се основните "градежни блокови" за синтеза на ткиво специфични протеини,…
Амино киселини се основните "градежни блокови" за синтеза на ткиво специфични протеини,…
 Образование лимфоцити прекурзори. Лезии на матични клетки
Образование лимфоцити прекурзори. Лезии на матични клетки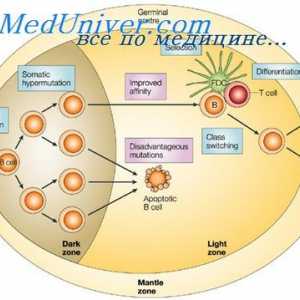 Плазма глувци тумори. Полирибозоми улога во биосинтезата на имуноглобулини
Плазма глувци тумори. Полирибозоми улога во биосинтезата на имуноглобулини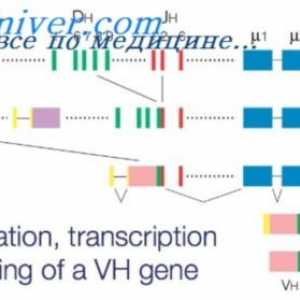 Ефектот на фенотип на висока синџирот на антитела. Ограничување V-гени
Ефектот на фенотип на висока синџирот на антитела. Ограничување V-гени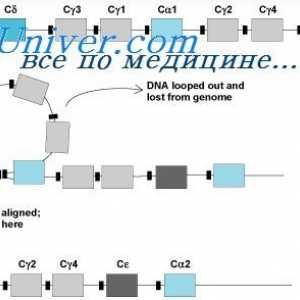 Методи за изолација на полирибозоми. Димензии полирибозоми синтетизираат антитела
Методи за изолација на полирибозоми. Димензии полирибозоми синтетизираат антитела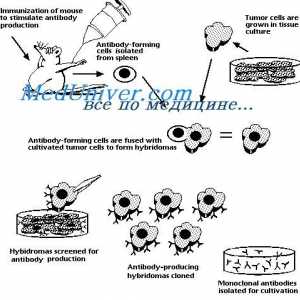 Превод на mRNA. Карактеристики на mRNA превод под синтеза на антителото
Превод на mRNA. Карактеристики на mRNA превод под синтеза на антителото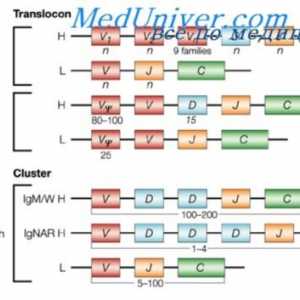 Translocon имуноглобулини. Комбинирање V- и гените на антитела
Translocon имуноглобулини. Комбинирање V- и гените на антитела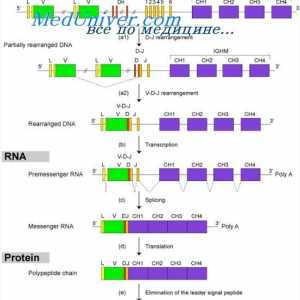 Формирање на имуноглобулин синџири. Вишокот антитело синтеза l-синџири
Формирање на имуноглобулин синџири. Вишокот антитело синтеза l-синџири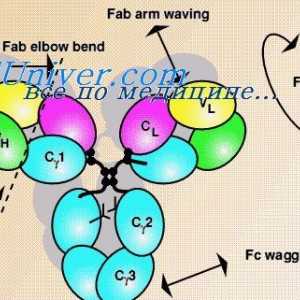 На варијабилни региони на антитела тешки ланци. Варијабилен регион гените на тешки ланци
На варијабилни региони на антитела тешки ланци. Варијабилен регион гените на тешки ланци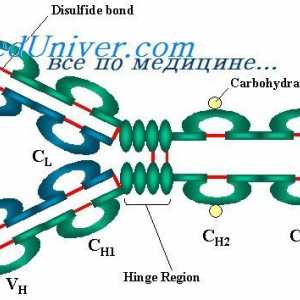 Имуноглобулин лесни ланци. организација на имуноглобулини
Имуноглобулин лесни ланци. организација на имуноглобулини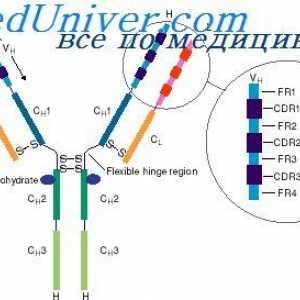 Локализација на гени лесни ланци антитело. Причините за променливоста на лесни ланци на
Локализација на гени лесни ланци антитело. Причините за променливоста на лесни ланци на