Translocon имуноглобулини. Комбинирање V- и гените на антитела
Во геномот на цицачите Постојат најмалку три различни translocon, од кои секоја е локализиран на различни nonhomologous autosomes. Во еден translocon содржеше V- и C-гени се утврди структурата на лесни ланци на капа-тип вториот - V- и C-ламбда светлина синџир гените-тип, и, конечно, трета translocon состои од V- и C-гени контролирање на формирање тешки ланци на сите класи и поткласи.
Комбинирање V- и C-гени во еден VC-транскрибирана ген се случува само во рамките на дадена translocon, при што на целиот сет на гени во рамките translocon станат активни само два гени - еден V и еден C разликуваат лимфоидни клетки од страна на имуноглобулин гени се врши не само во однос на тесно поврзани гени во во рамките на оваа translocon но исто така и против гени капа и ламбда nepey наоѓа на различни хромозоми.
Покрај тоа, за имуноглобулин гени познат феноменот на т.н. алелни исклучување, која се состои во фактот дека во одделни лимфоидни клетки е активно, само еден од двата алели (Mage, 1974).
Така, основните позиција клонална теорија избор на Burnet (Burnet, 1959), "една клетка - еден антитело" е реализиран од страна на фактот дека поединецот лимфоидни клетки од целиот сет на имуноглобулин гени активен само четири гени, имено VL, Cl и VH, CH.
На механизмите на активирање или репресија алелни гени и имуноглобулин гени, наоѓаат на различни autosomes, постојат сите податоци, и тука ќе се фокусираме на можните механизми на изборите "на" гени, кои сочинуваат една translocon.

Прв од сите таму е прашањето, на кое ниво постои поврзаност V- и C-гени во translocon. Во принцип, постојат три можности: Здружение на активноста на V- на производите и C-гени на ниво на полипептидот tsepi- асоцијација на ниво на mRNA транскрибирани одделно од V- и C-асоцијација на генот V- и C-гени во еден VC-ген на ниво на ДНК .
Постојат силни докази дека само Тоа е оваа претпоставка асоцијација V- и C-гени се случува на ниво на ДНК. На тој начин, што беше прикажано дека вклучувањето на радиоактивни амино киселина во процесот на преведување, и двете светлина и висока полипептидни ланци се случува рамномерно зголемување на должини синтетизираниот полипептид синџири и димензии полирибозоми да одговара на должината на mRNA, доволно за да еден молекул на mRNA е прочитан полипептид должина на синџир 450 амино киселини во случај на тешка возвиши и 210 амино киселини во случај на светлина синџир (Вилијамсон. 1971).
Изолирани од клетки глушец миелом mRNA способен за стимулирање на биосинтезата на комплетен полипептид синџир како систем клеточно-слободен под услови и во жаба ооцити.
РНК имуноглобулин лесни ланци, како и други протеини mRNA има поли А секвенца на 3`-крајот на молекулата. Нејзината молекуларна тежина е 400 000, што одговара до 1200 нуклеотиди. Околу 700 бази потребно е да се кодираат протеини себе има 200 бази на подот и A-сајт, и околу 300 база функција е непознат (Milstein д. A., 1972). Ако V- Унијата и C-гени се случува на ниво на ДНК, секвенца на бази во mRNA мора да биде континуирано и протеински секвенци одговараат на VC-граничната област.
Навистина, кога студија mRNA примарната структура глушец на белите дробови синџири даде дека v има база на секвенца на mRNA што одговара на амино киселина остатоци од 105- 108 на лесниот ланец, односно. e. граница на делот помеѓу V-и-пол коло.
друг доказ за асоцијација V- и C-гени на ниво на ДНК се студии, во кои се изучува структурата на протеините во т.н. тешки ланци на човечките болести. Во оваа болест, со тешки ланци се скратува со бришење на регионите на синџирот на полипептид, кои во многу случаи е вклучена `низа кој се протега на границата меѓу дел на променливата и постојано региони на колото.
А слична бришење експериментално успеа во добивање на анализа на клетки мутанти глушец миелом (Milstein e. a., 1974), а во вториот случај, тоа е познато дека делеција се случи по комбинирање на V- и C-гени, што како почетен клеточна линија синтетизира со тежок ланец на нормална должина.
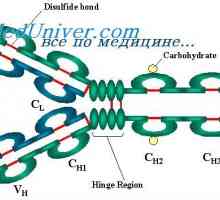 Имуноглобулин лесни ланци. организација на имуноглобулини
Имуноглобулин лесни ланци. организација на имуноглобулини Контрола на образование имуноглобулин лесни ланци. Гените лесни ланци антитело
Контрола на образование имуноглобулин лесни ланци. Гените лесни ланци антитело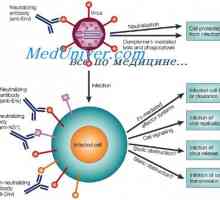 На антигеноста на имуноглобулини. Антигенски детерминанти на антитела
На антигеноста на имуноглобулини. Антигенски детерминанти на антитела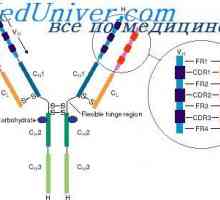 Локализација на гени лесни ланци антитело. Причините за променливоста на лесни ланци на
Локализација на гени лесни ланци антитело. Причините за променливоста на лесни ланци на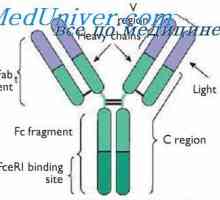 На варијабилни региони на светлината антитело синџири. Гени променлив регион
На варијабилни региони на светлината антитело синџири. Гени променлив регион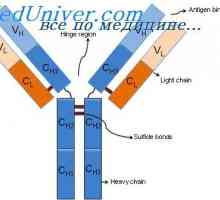 Алелни варијанти на имуноглобулин g. Гените на тешки ланци IgG
Алелни варијанти на имуноглобулин g. Гените на тешки ланци IgG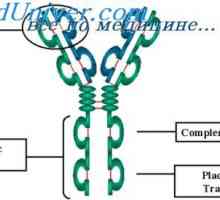 Cast лесни ланци на имуноглобулин. Ekstrauchastki антитела
Cast лесни ланци на имуноглобулин. Ekstrauchastki антитела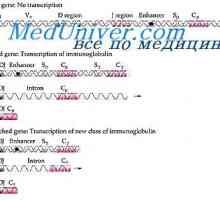 Формирање VC-имуноглобулинот ген. Механизми асоцијација V- и гените на антитела
Формирање VC-имуноглобулинот ген. Механизми асоцијација V- и гените на антитела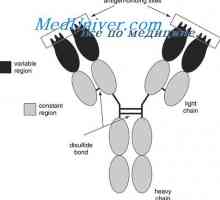 Образование имуноглобулин тежок ланец. Константен регион со тежок синџир на антителото
Образование имуноглобулин тежок ланец. Константен регион со тежок синџир на антителото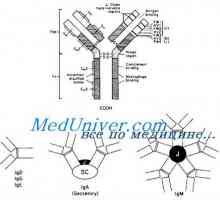 Заплеткувањето антитело со тежок ланец гени. Гените на тешки ланци IgA
Заплеткувањето антитело со тежок ланец гени. Гените на тешки ланци IgA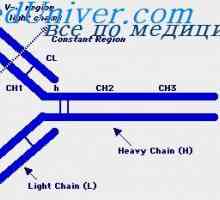 Прилог VH имуноглобулин гени. Хромозом со гени антитело
Прилог VH имуноглобулин гени. Хромозом со гени антитело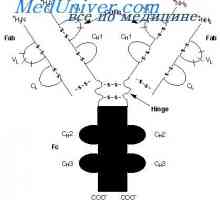 Разликите варијабилни региони на антитела. Променлив подгрупа имуноглобулин
Разликите варијабилни региони на антитела. Променлив подгрупа имуноглобулин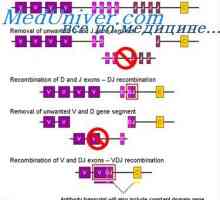 Уредување VC-ген. Прекинувачки антитело ген активност
Уредување VC-ген. Прекинувачки антитело ген активност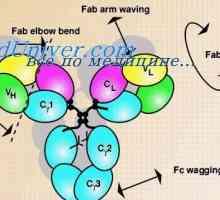 На варијабилни региони на антитела тешки ланци. Варијабилен регион гените на тешки ланци
На варијабилни региони на антитела тешки ланци. Варијабилен регион гените на тешки ланци Формирање на имуноглобулин синџири. Вишокот антитело синтеза l-синџири
Формирање на имуноглобулин синџири. Вишокот антитело синтеза l-синџири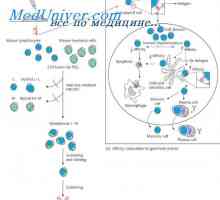 Гените синтетизирање на антитело. Бројот на гени кои се вклучени во синтезата на имуноглобулини
Гените синтетизирање на антитело. Бројот на гени кои се вклучени во синтезата на имуноглобулини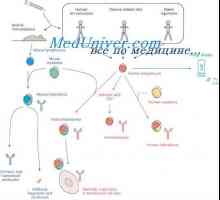 Хибридизација на mRNA и ДНК антитела. Локализација V- и гени во геномот на имуноглобулини
Хибридизација на mRNA и ДНК антитела. Локализација V- и гени во геномот на имуноглобулини Симултан синтеза на различни имуноглобулини. Хематопоетски матични клетки
Симултан синтеза на различни имуноглобулини. Хематопоетски матични клетки Ефектот на фенотипот на генот активност. Алелни детерминанти на антитела
Ефектот на фенотипот на генот активност. Алелни детерминанти на антитела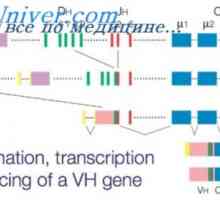 Ефектот на фенотип на висока синџирот на антитела. Ограничување V-гени
Ефектот на фенотип на висока синџирот на антитела. Ограничување V-гени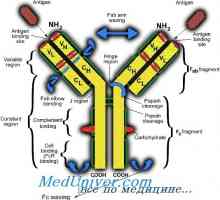 Хипогамаглобулинемија кај децата. Недостаток на имуноглобулин G (IgG) детето
Хипогамаглобулинемија кај децата. Недостаток на имуноглобулин G (IgG) детето
 Симултан синтеза на различни имуноглобулини. Хематопоетски матични клетки
Симултан синтеза на различни имуноглобулини. Хематопоетски матични клетки Хипогамаглобулинемија кај децата. Недостаток на имуноглобулин G (IgG) детето
Хипогамаглобулинемија кај децата. Недостаток на имуноглобулин G (IgG) детето Формирање VC-имуноглобулинот ген. Механизми асоцијација V- и гените на антитела
Формирање VC-имуноглобулинот ген. Механизми асоцијација V- и гените на антитела Образование имуноглобулин тежок ланец. Константен регион со тежок синџир на антителото
Образование имуноглобулин тежок ланец. Константен регион со тежок синџир на антителото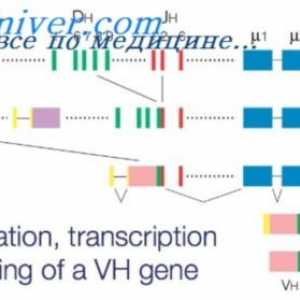 Ефектот на фенотип на висока синџирот на антитела. Ограничување V-гени
Ефектот на фенотип на висока синџирот на антитела. Ограничување V-гени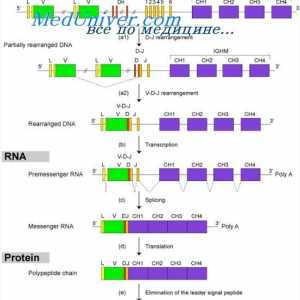 Формирање на имуноглобулин синџири. Вишокот антитело синтеза l-синџири
Формирање на имуноглобулин синџири. Вишокот антитело синтеза l-синџири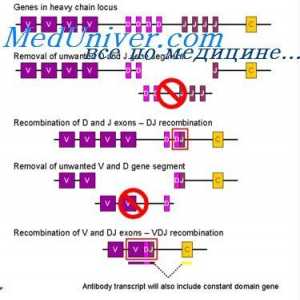 Уредување VC-ген. Прекинувачки антитело ген активност
Уредување VC-ген. Прекинувачки антитело ген активност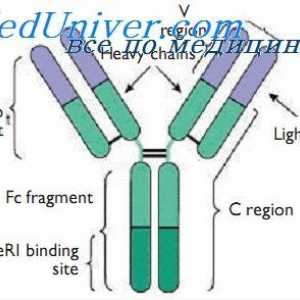 На варијабилни региони на светлината антитело синџири. Гени променлив регион
На варијабилни региони на светлината антитело синџири. Гени променлив регион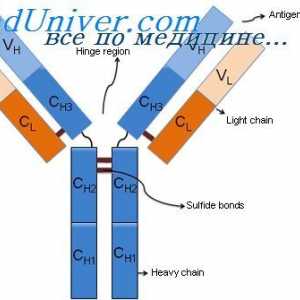 Алелни варијанти на имуноглобулин g. Гените на тешки ланци IgG
Алелни варијанти на имуноглобулин g. Гените на тешки ланци IgG На варијабилни региони на антитела тешки ланци. Варијабилен регион гените на тешки ланци
На варијабилни региони на антитела тешки ланци. Варијабилен регион гените на тешки ланци