Неврофизиолошки механизми за побудување на визуелен пат

Како дел од оптичкиот нерв аксони од повеќето од ретинална ганглион клетките влегува во странични геникулатно тело.
Некои аксони се испраќаат до tectal и pretectal области на мозокот. проекции Retinopretektalnye имаат retinotopichekuyu организација. Информациите доставени до tectum е вклучена во регулирањето на движењето на очите и пупиларните реакции. Дел од аксони на ретинална ганглион клетките е проектиран во три јадра на хипоталамусот, перница и дополнителни средината на мозокот јадра. Нервни влакна од ретинална ганглион клетките, достигнувајќи хипоталамусот, очигледно се анатомска подлога за контрола на светло на деноноќниот ритам. Влакна кои доаѓаат за да го ублажат таламусот се на системот за окуломоторен, а влакната се вклучени во дополнителни јадра може да учествува во визуелната проприоцепција обезбедување стабилизација на визуелна слика на мрежницата.
Во хијазма на оптички влакна кој се протега од ретината на двете очи на BWL, се сечат. Затоа, оптички тракт и внесете ја тестира секоја BWL оптички влакна од временската половина ипсилатерални ретината и половина од носната контралатерални мрежницата. Повеќето од аксони на оптичкиот тракт завршува во ипсилатерални LKT. А мал пакет на нервни влакна се протега помеѓу два лобуси на ипсилатерални супраоптичките јадро и паравентрикуларните јадро се навраќа на хипоталамусот. Можеби овие нервни влакна се нервни влезови за контрола на деноноќниот ритам.
Секоја група на влакната на оптички тракт е испратен на медијална геникулатно тело, формирајќи комисура Gudden. Функцијата на овие влакна е нејасно. Еден значаен дел од форми оптички тракт синаптичките затворање pretectal јадро во средниот мозок и се дел од лак pupillomotornogo светлина рефлекс.
Трансфер на возбудувањето и ширење на пулс по должината на оптичкиот пат имаат свои карактеристики. Оптичкиот нерв влакна покриени од страна на миелинската обвивка. миелинската обвивка на нервните влакна има повисока отпорност (500-800 мегаоми / cm2) И ја врши функцијата на изолатор, спречување на загуба на струја во нервните влакна помеѓу јазли на Ranvier. Исто така, за миелинската обвивка мала количина карактеристика на одредени капацитет, што се должи на значително дебелина и добри диелектрични својства на миелинската обвивка.
Благодарение на овие својства, миелинизираните нервни влакна на оптичкиот нерв потенцијали акција спроведена екстремно брзо. Само за многу кратко делови од овие влакна немаат миелинската обвивка и се покриени со конвенционалните клеточната мембрана (јазли на Ranvier). Ширењето на побудување во нервни влакна не се случува постојано, но нередовните, односно. Е. Saltatory. Доцнењето на пулсот може да биде само на јазли на Ranvier.
Брзината на интензитет по должината на аксоните на ретинална ганглион клетките, односно. Д. оптички пат зависи од дијаметарот на секоја нервните влакна. Со релативно дебели миелинизирани аксони на ретинална ганглион клетки на стапката на побудување е висока - 35-50 m / s (Y-неврони), на тенок миелинизирани аксони на брзината е 15-25 m s (X неврони) и слабо миелинизирани аксони брзина / на уште помала - 5-9 m / s.
Така, висока брзина на пулсот во миелинизираните нервни влакна на оптичкиот нерв дава можност на постоење на голем број на паралелни bystroprovodyaschih нервни патишта. Кога снимање на визуелни евоцирани потенцијали од здрава личност пулсот на флеш светлина се пренесува по должината на оптичкиот пат 70 ms, со што максимален одѕив на визуелниот кортекс во просек веќе по 100 ms (P бран100).
Во демиелинизирачки заболувања на нервниот систем во кој нервните влакна на оптичкиот пат изгуби миелинската обвивка пулсот протокот по должината на оптичкиот пат се забави или целосно запрен. Затоа кортикални време (чекање пулсот на ретината фоторецептори да лае) и retinokortikalnoe време (држи пулсот од ретинална ганглион клетките на кортексот) се дијагностички критериуми патологија визуелен пат и се користат во клиничката пракса за рана дијагноза на невритис, исхемија, мултиплекс склероза и атрофија на оптичкиот нерв.
} {Модул direkt4
Исто така, аксонална невропатија забележани во која скршени на аксонот (axoplasmatic) превоз по должината на нервните влакна.
Трансфер на побудување од неврон на неврон синапсите се случува преку. Нервните импулси достигнува аксонот анатомски крајот и предизвикува ослободување на специфични невротрансмитер молекули во синаптичката пукнатина. Молекула невротрансмитери многу протеини помали молекули, но поголема од натриум или калциум јони. Во моментов тоа одвои повеќе од 20 хемиски медијатори во мрежницата: ацетилхолин, норадреналин, допамин, глицин, глутамат, аспартат, serogonin, гама-аминобутерна киселина (ГАБА), итн кога посредниците се ослободени од пресинаптичката аксон мембрана, тие брзо се дифузни низ синаптичката пукнатина да. постсинаптичката мембрана од следниот неврон.
Постсинаптичката мембрана е, исто така, функционално специјализирани: тоа содржи протеини рецептори, кои реагираат на невротрансмитерот соодветните отворање на јонските канали, преку која на различни јони. Пред очите кој поминува низ мембрана јон на постсинаптичка (натриум, калиум, хлориди и други.) Во зависност деполаризација на потенцијалот на невронот мембрана или стабилизација.
Така, на нервните импулси оди до крајот на аксонот и предизвикува тука специфични молекули ослободување невротрансмитер кој делува на постсинаптичката мембрана, или да се спречи нејзиното намалување (стабилизирајќи). фреквенцијата на пулсот се зголемува со намалување на мембранскиот потенцијал. Ова стимулирачки синапси. Ако потенцијал на мембраната се стабилизира на ниво на под-праг, фреквенцијата на пулсот се намалува, или тие не се случи. Ова инхибиторни синапсите. Разлика дали активно или сопирање стимулира синапса - зависи од видот на ослободен во него невротрансмитер рецептор молекули и на постсинаптичката мембрана.
Норепинефрин - стимулирачки невротрансмитер GAM К - сопирање. Функционална специјализација на секој синапсите е константна во текот на животот.
А многу комплицирана структура пресинаптичката оска терминали на нервните клетки и постсинаптичката мембрана од следниот невронот предизвикува само мало доцнење на потенцијалните брзина. студии морфометрички покажаа дека постојат два типа на промени: намалување должина сооднос mezhperehvatnogo дел со дијаметар нервните влакна (L / D) 10-15 (во миелинизираните аксони, оваа вредност е обично 150-200) и намалување на дијаметарот на аксијален цилиндар на нервните влакна ( d).
Magnons и parvocellular начин визуелен систем. Визуелен систем има паралелни патеки кои ги поврзуваат мрежницата на визуелна центри на низводно. Во исто време, постои јасна retinopaticheskaya организација паралелно аферентните врски на мрежницата.
Странични геникулатно тело добива голем дел од патот на оптички влакна. Дел аксони BWL поминува и завршува во други структури на диенцефалонот и средниот мозок (супериорен коликулус, pretectal област, гуми јадро).
Во функционална смисла, најважните два нервни патишта: magnocellular (М) патека и parvocellular (P-патека). Овие патеки - главните информации канали на визуелниот систем се протега од мрежницата на LKT и примарниот визуелен кортекс (зона VI).
Во мрежницата на М-ганглија клетки се голем чадор клетки кои имаат големи мобилен тела, дебели аксони и богат дендрично разгранување. P-ретинална ганглион клетките имаат помали мобилен тела, аксони, и мали тенки дендрично разгранување, но со густа разгранување. Тие вклучуваат midzhitganglioznye клетки. Меѓу ретинална ганглион клетките и М-Р-клетки го сочинуваат мнозинството, 10% и 80% од населението, соодветно. M- и P-ганглиските клетки наоѓа во близина на едни со други во ганглиски клетки слој, е формирана извртени но независно мозаик низ мрежницата.
странични геникулатно тело, исто така, има специфичноста на анатомски и функционални организација, како одраз на постоењето на два паралелни визуелни системи за обработка.
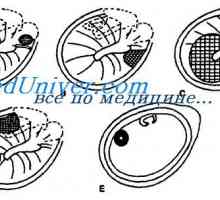 Оптичкиот нерв на ембрионот. овошје окуломоториус нерв
Оптичкиот нерв на ембрионот. овошје окуломоториус нерв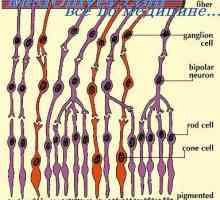 Histogenesis ембрионски мрежницата. Морфологијата на фетусот ретинална
Histogenesis ембрионски мрежницата. Морфологијата на фетусот ретинална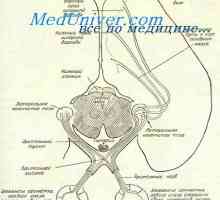 Формирањето на нервните ембрионот око. Патишта на визуелна центри на фетусот
Формирањето на нервните ембрионот око. Патишта на визуелна центри на фетусот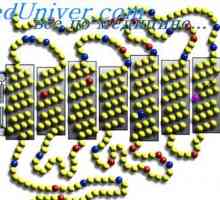 Amacrine ќелија. Ретинална ганглион клетките
Amacrine ќелија. Ретинална ганглион клетките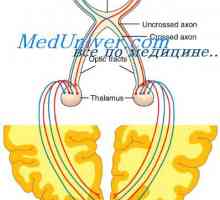 Визуелен начин. Дорзолатерално геникулатно јадро на таламусот
Визуелен начин. Дорзолатерално геникулатно јадро на таламусот Анатомски и топографски карактеристики и функции на оптичкиот нерв
Анатомски и топографски карактеристики и функции на оптичкиот нерв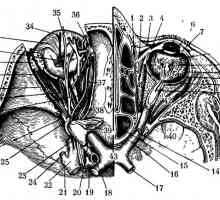 Структурата на оптичкиот нерв
Структурата на оптичкиот нерв- Дијаметарот на ученик
 Кернелот патека на визуелните анализатор. Јадрата на гледање. Знаци на оптичкиот тракт.
Кернелот патека на визуелните анализатор. Јадрата на гледање. Знаци на оптичкиот тракт.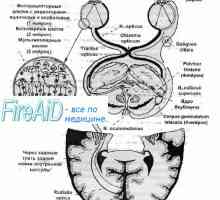 Патишта. Патека на визуелните анализатор. проводен поглед на патот.
Патишта. Патека на визуелните анализатор. проводен поглед на патот.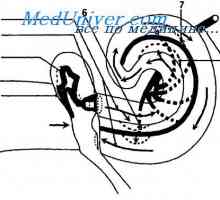 Патеката на звук бран. Аферентните инервација на кохлеата.
Патеката на звук бран. Аферентните инервација на кохлеата.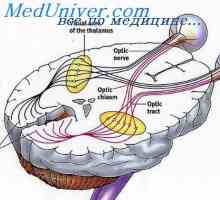 Анатомија на визуелен пат. Централна неврони на визуелен пат
Анатомија на визуелен пат. Централна неврони на визуелен пат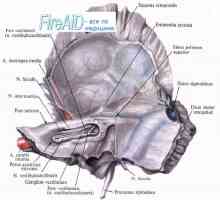 Аудитивни нерв. Невролошки патот на аудитивни нерв
Аудитивни нерв. Невролошки патот на аудитивни нерв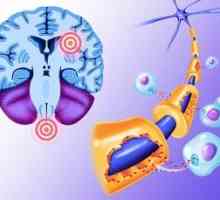 Патогенетски механизми на нарушување на визуелната функција кај пациенти со мултиплекс склероза
Патогенетски механизми на нарушување на визуелната функција кај пациенти со мултиплекс склероза Анализа на визуелните сигнали во примарната зона (стријатна) визуелен кортекс
Анализа на визуелните сигнали во примарната зона (стријатна) визуелен кортекс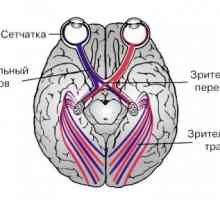 Анатомски топографски карактеристики на визуелен пат
Анатомски топографски карактеристики на визуелен пат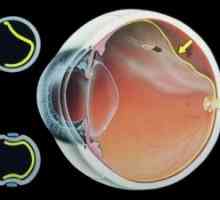 Ембриогенезата мрежницата
Ембриогенезата мрежницата Ембриогенезата, оптичкиот нерв
Ембриогенезата, оптичкиот нерв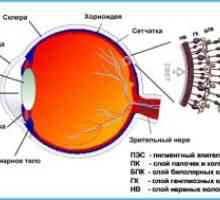 Еволуцијата на сензорни функција на ретината во текот на ембриогенезата и раните години
Еволуцијата на сензорни функција на ретината во текот на ембриогенезата и раните години Миелинизација на нервните влакна на оптичкиот пат
Миелинизација на нервните влакна на оптичкиот пат Периферните и централните делови на нервниот начин pupillomotornogo
Периферните и централните делови на нервниот начин pupillomotornogo
 Ембриогенезата мрежницата
Ембриогенезата мрежницата Патогенетски механизми на нарушување на визуелната функција кај пациенти со мултиплекс склероза
Патогенетски механизми на нарушување на визуелната функција кај пациенти со мултиплекс склероза Кернелот патека на визуелните анализатор. Јадрата на гледање. Знаци на оптичкиот тракт.
Кернелот патека на визуелните анализатор. Јадрата на гледање. Знаци на оптичкиот тракт. Анализа на визуелните сигнали во примарната зона (стријатна) визуелен кортекс
Анализа на визуелните сигнали во примарната зона (стријатна) визуелен кортекс Еволуцијата на сензорни функција на ретината во текот на ембриогенезата и раните години
Еволуцијата на сензорни функција на ретината во текот на ембриогенезата и раните години Миелинизација на нервните влакна на оптичкиот пат
Миелинизација на нервните влакна на оптичкиот пат Amacrine ќелија. Ретинална ганглион клетките
Amacrine ќелија. Ретинална ганглион клетките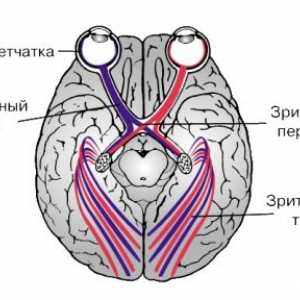 Анатомски топографски карактеристики на визуелен пат
Анатомски топографски карактеристики на визуелен пат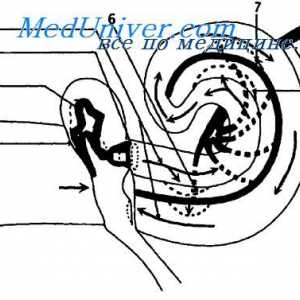 Патеката на звук бран. Аферентните инервација на кохлеата.
Патеката на звук бран. Аферентните инервација на кохлеата.