Дефект на убиквитин-протеазом патека протеин
Повеќето (80-90%) интрацелуларен протеин дефект врши убиквитин-протеазом патека. патека на убиквитин-протеазом е присутна во јадрото и цитоплазмата на еукариотските клетки и игра важна улога во разградувањето на нормални и абнормални протеини. Овој пат е одговорен за контролирано деградација на многу протеини, вклучувајќи ги и оние кои се потребни за контрола на растот и размножувањето на клетките, клеточната диференцијација, имунолошки и воспалителни реакции, апоптоза и метаболни адаптација.
патека убиквитин-протеазом исто така, врши "економски" функција главно од протеини циркулација и елиминација на абнормално протеини со погрешна кодирање, misfolded локализиран во не-основни области, оштетени или се преземени во спротивно надвор од акција. патека на убиквитин-протеазом игра клучна улога во контролата на мускулната маса, а неговата активност е зголемена во кахексија. Овој пат, исто така, игра значајна улога во обновувањето на мускулите и преуредување.
патека убиквитин-протеазом Тоа може да се гледа како секвенца од три процеси:
(1) препознавање на протеини супстрат за да се распаѓање;
(2) ковалентна прикачување на polyubiquitin ланци како протеин ознака за распаѓање;
(3) протеолиза 2500 kD протеински комплекс, наречен протеазом 26S.
препознавање на протеини, наменети за распаѓање обично се врши:
(1) според некои структурни промени на протеини, вклучувајќи ги и ефектите на секвенца на специфична амино киселина која е скриена во нормален пост-транслациони модификации, како што фосфорилација или хидроксилација;
(2) за спојување или ослободува својот лиганди;
(3) интеракција со адаптер протеин или придружникот (на пример, извоз misfolded протеини од страна на chaperones во цитозолот на ИПР);
(4) специфични штета се случуваат во протеин по пат на оксидација или nitrosylation.
Покрај тоа, присуството на одредени "дестабилизирачки"Остатоци IV-end дел на пептид ознаката наменета за расцеп (на пример, со краток полу-живот). Сепак, тоа треба да се напомене дека пред да биде подложен на поделба, не сите протеини произведени убиквитин таг.
Спротивно на тоа, некои протеини се расцеп од 20-тите години големи протеазом. Целта на овој модел е јасно разделување, но се чини дека се случува со протеини имаат јасно неструктурирани региони, давајќи им можност на протеини поголема нестабилност.

Првата реакција - активирање на убиквитин E1 (ubikvitinaktiviruyuschim ензим), а потоа доставени до убиквитин E2 (ubikvitinprisoedinya yuschemu-ензим).
На вториот реакција - формирање на комплекс со E2, EZ (убиквитин поврзување на ензим) и подлогата.
На третата реакција - трансфер на убиквитин лизин (-AM) супстрат, подлогата за да се одбележи polyubiquitin синџир. Како резултат на тоа, четвртиот чекор е одвоена од подлогата polyubiquitin Е3.
Конечно, супстрат на протеазом unrolls за ATP-зависни начин од страна на активноста на ubikvitingidralazy одвојува убиквитин синџирот и внесува ширевте протеин во комората за протеазомот се делови каде протеаза активност. Молекулите на убиквитин се рециклира, а како резултат на пептидите се расчленети на амино киселини и користи повторно за синтеза на нови протеини.
УБ - убиквитин.
По протеини беше идентификуван како супстрат до деградација, тоа е предвидено со ковалентно етикетирани убиквитин. Убиквитин - протеини присутни во сите типови на клетки и се состои од остатоци од 76 амино киселина вклучувајќи C-терминалниот глицин и лизин остаток на позиција 48. Убиквитин е ковалентно врзан за протеинот наменета за деградација во серија од три реакции катализирана од ензими познат како E1 (ubikvitinaktiviruyuschy ензим), Е2 (ubikvitinprisoedinyayuschy ензим) и Е3 (ubikvitinsvyazyvayuschy ензим).
таму Две изоформи Е1, Неколку изоформи на Е2 и Е3 е многу голем број на ензими, овозможувајќи за специфични супстрати на многу ткива и регулирање на овој процес.
првата молекула убиквитин активира од страна на врзување на ATP-зависни реакција на Е1, а потоа префрлен на убиквитин E2 фрагмент. И Е1 и Е2 се активни сајтови на цистеин остатоци кои ја формираат thioesters со C-терминалниот глицин остаток од убиквитин. Конечно, убиквитин приклучи на Е2 се пренесува директно или преку внатрешниот Е3 lizilovomu остатоци на протеини супстрат.
Е3 игра важна улога во признавањето протеин супстрат за расцепување и во регулирањето на формирање на подлогата комплекс E2 / Е3. На сличен начин понатаму убиквитин молекули се прикачени за супстрат кој има прикачен убиквитин молекула, формирајќи isopeptide врски помеѓу C-терминалниот глицин остаток од убиквитин молекула и лизин наоѓа на позиција 48 пред додаде убиквитин молекула.
потребни синџирот на најмалку четири молекули на убиквитин на протеини неколку молекули означени убиквитин беа лесно препознатливи и се насочени кон 26S протеазоми расцеп. Неодамна, една анкета била обврзувачка области на активност и убиквитин. Поважно, процесот на ubiquitination е реверзибилна, со присуство на процесот на исклучување, која се јавува класа на цистеин протеази нарекува deubikvitiniruyuschimi ензими.
активност deubikvitiniruyuschego ензим Тоа е да се елиминираат врзување на убиквитин, убиквитин обработка претходник, поврзувајќи убиквитин синџири и повторната употреба на убиквитин. Овие процеси се одговорни за регулирање на неколку сигнални патишта кои се важни за процесот на развој, вклучувајќи ги и клеточниот раст и диференцијација.
вистинските деградација на ubiquitinated протеини се случува во внатрешноста на комората на протеазом, но на молекули на убиквитин одвојуваат првиот, така што тие може да се користи, на протеазом 26S е голем, која се состои од неколку подединици на комплексот, која ги вклучува субединица 20-ти како протеолитички јадро со регулаторни 19s комплекс прикачена на една или двата краја. Регулаторни субединици се вклучени во препознавање на означените протеини, отстранување убиквитин ознака, како и на ATP-зависни процеси ваквиот развој на протеинот и неговите протеолитички основни во еден правец има форма на тунел.
протеолитички 20-ти комплекс е буре-како структура која се состои од четири наредени една врз друга прстенест структури (авва), од кои секоја е формирана од страна на седум подединици. На централната празнина на каталитички структура се состои од вкупно шест протеолитички делови формирана три различни каталитички подединици секое b-прстен. Тие се однесуваат на каталитичка субединици ТВ-терминал треонин gidralazam како IV-терминал треонин делува како нуклеофилна катализатор. Сепак, три различни подединици во секој од двата прстени се различни параметри во расцепување на пептидни врски веднаш по основните, хидрофобни и киселински остатоци.
20 подгрупата хидролизира дојдовни супстрат во пептидни фрагменти која се состои од 3-30 аминокиселински остатоци. Овие пептид производи се ослободени од протеазомот во клетка, а потоа се подложат на хидролиза други протеази и aminopeptidases.
регулатива протеолиза во протеазом се јавува во три нивоа. Прво, препознавање на подлогата е прилагодлив функции кои го за цел протеин poliubikvitinirovaniya на. За повеќето протеини се неидентификувани подлоги и вклучуваат фосфорилација, хидроксилација на пролин резидуа или идентификување расцеп сигнал содржани во основното низа. Второ, регулирани деградација на специфичните класи на супстрати може да се постигне со комбинирање на комплекси E2 / ЕЗ со разни помошни фактори.
На пример, во некои случаи, имено EZ Тоа мора да се промени или "вклучен" низ посттранслациона модификација да преземе активна форма која го признава супстрат. Во други случаи, на стабилноста на протеини подлогата зависи од неговата поврзаност со молекуларна chaperones, кои дејствуваат како елементи откривање и кои служат како врски со соодветни ligases. Во особено, инсулин се намалува убиквитин-протеазом активност со посредство на интрацелуларни протеази и движење insulinosnizhayuschego ензим од 20-тите години на протеазом и 26S. Конечно, патека на убиквитин-протеазом може да се контролира од страна на било интеракција или промени во изразувањето на убиквитин протеазом субединици.
Еден пример за ова може да се забележи по Интравенозна инфузија на амино киселини или по нивното воведување во цревниот лумен. Зголемување на испорака на амино киселини, но не гликоза намалува mRNA изразување на убиквитин ubikvitinprisoedinyayuschego ензим со молекуларна тежина од 14 kDa, и C9 протеазом субединица во интестиналната мукоза.
 Сигнализација патека од малиот мозок. Пуркиниевите клетки на малиот мозок
Сигнализација патека од малиот мозок. Пуркиниевите клетки на малиот мозок На разградувањето на протеините. Физиологија дефект регулирање протеин
На разградувањето на протеините. Физиологија дефект регулирање протеин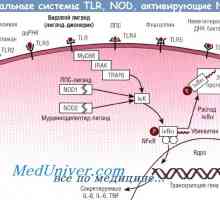 Embodiments obrazraspoznayuschih prrs рецептори. Toll-како рецептори
Embodiments obrazraspoznayuschih prrs рецептори. Toll-како рецептори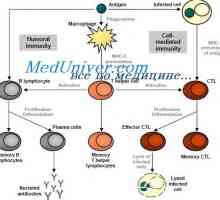 Сигнали патарина-како рецептори. Улогата на молекулата myd88
Сигнали патарина-како рецептори. Улогата на молекулата myd88 Намалување на ризикот од алергии со помош на инфекција. Имунолошкиот одговор на вирусот херпес…
Намалување на ризикот од алергии со помош на инфекција. Имунолошкиот одговор на вирусот херпес…- Ефект на бактерии во цревата на својата инфламаторен одговор
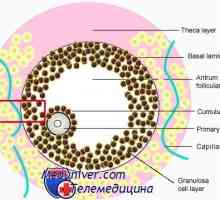 Знаци на подготвеност за ооцитот мејозата. Распуштање зародишен везикули (m)
Знаци на подготвеност за ооцитот мејозата. Распуштање зародишен везикули (m) Состојбата на цревните епителни имунитет кај недоносените
Состојбата на цревните епителни имунитет кај недоносените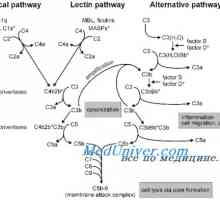 Класичната патека на активацијата на комплементот
Класичната патека на активацијата на комплементот Механизми на забите и катаболизам на протеините во организмот
Механизми на забите и катаболизам на протеините во организмот Не-геномната ефекти на стероиди. G-протеин-спарен рецептори
Не-геномната ефекти на стероиди. G-протеин-спарен рецептори Патогенеза начин NFkB / rel во реакција на бактерии од интестиналниот епител
Патогенеза начин NFkB / rel во реакција на бактерии од интестиналниот епител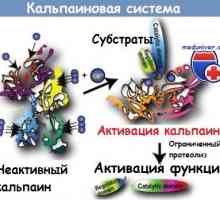 Kalpainovaya систем протеолиза
Kalpainovaya систем протеолиза Autofagalno-лизозомални систем на апоптоза протеин расцепување
Autofagalno-лизозомални систем на апоптоза протеин расцепување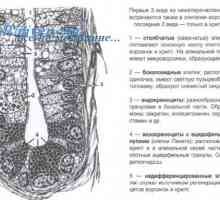 Улогата на сигнализација патека протеини морфогенеза на коските (BMP) во диференцијација на матични…
Улогата на сигнализација патека протеини морфогенеза на коските (BMP) во диференцијација на матични…- Значење WNT сигнализација патека во регулирање на цревната матични клетки
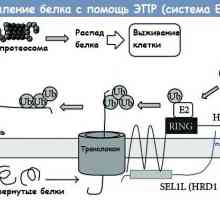 Расцепување на протеини преку ендоплазматичниот ретикулум (ER) erad систем
Расцепување на протеини преку ендоплазматичниот ретикулум (ER) erad систем Создаден ветувачки лек за рак на бубрезите
Создаден ветувачки лек за рак на бубрезите- Антиоксиданс Брокулата може да се третира progeria?
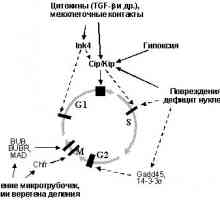 Onkologiya-
Onkologiya-- Недостаток на цинк и нашите болки
 Состојбата на цревните епителни имунитет кај недоносените
Состојбата на цревните епителни имунитет кај недоносените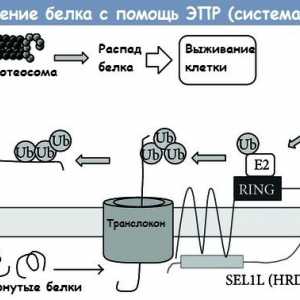 Расцепување на протеини преку ендоплазматичниот ретикулум (ER) erad систем
Расцепување на протеини преку ендоплазматичниот ретикулум (ER) erad систем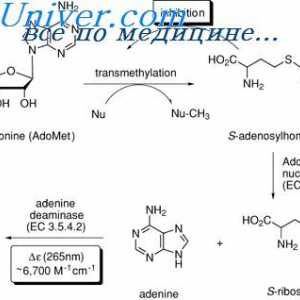 На разградувањето на протеините. Физиологија дефект регулирање протеин
На разградувањето на протеините. Физиологија дефект регулирање протеин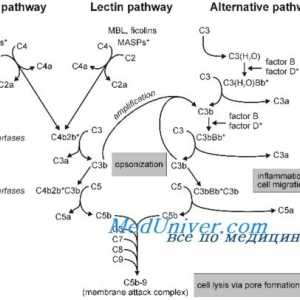 Класичната патека на активацијата на комплементот
Класичната патека на активацијата на комплементот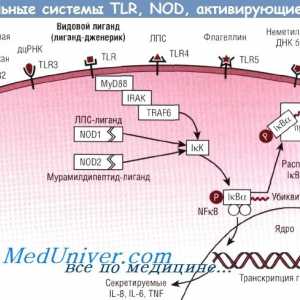 Embodiments obrazraspoznayuschih prrs рецептори. Toll-како рецептори
Embodiments obrazraspoznayuschih prrs рецептори. Toll-како рецептори Намалување на ризикот од алергии со помош на инфекција. Имунолошкиот одговор на вирусот херпес…
Намалување на ризикот од алергии со помош на инфекција. Имунолошкиот одговор на вирусот херпес…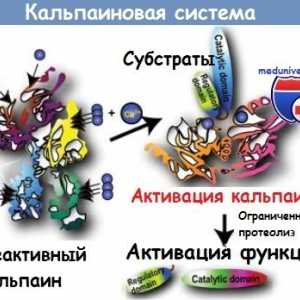 Kalpainovaya систем протеолиза
Kalpainovaya систем протеолиза