Рецептори за хормони поврзани со G-протеин. ензим-конјугиран хормонски рецептори на
Видео: Студија Г протеини. Stand-up од лабораторија
многу хормони активирани рецептори кои се регулира активноста на протеини, целни клетки (на пример, ензими или јонски канал протеини) не се директно, но со комбинирање групи со клеточната мембрана протеин наречен heterotrimeric GTP-врзани протеини (G-протеини). 1000 познати G-протеин-спарен рецептори, тие имаат 7 трансмембрански сегменти формирање на јамки во рамките на и надвор од клеточната мембрана. Некои делови на рецепторот, кои дејствуваат во цитоплазмата на клетките (главно се наоѓа во цитоплазмата на опашката на клеточниот рецептор е), во комбинација со Г-протеини, вклучувајќи ги и трите делови (со тоа - тример): алфа, бета, гама-подединици. Кога лиганд (хормон) се поврзува со екстрацелуларниот дел на рецепторот во него се подложат на конформациона промени што ги активираат G-протеин и интрацелуларни сигналите што го индуцираат кои се или (1) отворање или затворање на мембрана јонски канали, или (2) да се промени на активноста на ензимите во цитоплазмата.
Три-G-протеини Го добила своето име, поради неговата способност да се поврзе со гванозин нуклеотиди. Во неактивна состојба на алфа, бета, гама-субединица на Г-протеин се формира комплекс во врска со гванозин дифосфат од страна на алфа подединици. Кога е активиран рецептор, последователните конформациона промени доведуваат до замена на гванозин дифосфат и гванозин трифосфат во цитоплазматски дел на рецепторот интеракција со Г-протеин. Замена на GTP на БДП води-субединица одделени од три-компонента комплекс и во интеракција со други мобилен сигнализација протеин. Овие протеини, од своја страна, да ја смени активност или јонски канал, клетки или ензими, како што аденилат циклаза и фосфолипаза C, промена на функцијата на клетката.
докази активирање исчезнат поради исчезнувањето gormona- алфа подгрупата се инактивира со замена на Асошиетед GTP на БДП. Потоа, A-субединица на ново комбинација бета и гама-субединици за да се формира неактивни тројна мембрана врзани G-протеин.
некои хормони интеракција со инхибиторни Г-протеини (назначено како G1-протеини), а од друга - со активирање на Г-протеини (означена како GS-протеини). Така, во зависност на рецепторот на интеракција на хормон со инхибиторен или активирање на G-протеински хормон или може да се зголемување или намалување на активноста на ензимите во клетката. Овој сложен систем на клеточната мембрана Г-протеини и потенцијално обезбедува спектар на различни одговори на целните ткива на дејството на различни хормони.

ензим-конјугиран хормонски рецептори на
некои рецептори, се активира за да функционираат како ензими, или се тесно поврзани со ензими кои се активирани. Овие ензимски-конјугиран рецептори се протеини кои имаат само еден трансмембрански дел, за разлика од G-протеин-спарен рецептори има седум такви фрагменти. Ензимско-конјугиран рецептори имаат хормон кој што врзува дел на површината на надворешната мембрана и го катализира или се поврзани со ензимот дел од внатрешната површина. Кога хормон се врзува за надворешниот дел од рецепторот, ензим кој е веднаш активирана (или деактивира), иако многу ензимски спарени рецептори имаат свои ензимската активност, додека други влијаат на ензими кои се тесно поврзани со рецепторот, која е придружена со промени во ќелија функции.
еден пример ензим-рецептор Коњугат може да биде на рецептор на лептин. Лептин е хормон кој се секретира масните клетки со директно, и предизвикува различни физиолошки ефекти. Меѓу особено важно неговите влијанија вклучуваат влијание врз апетитот и енергетскиот биланс.
рецептор лептин Тој е член на големото семејство на цитокински рецептори кои не се поседуваат ензимската активност, но нивната активност е во спрега со ензим сигнал. Во случај на рецептор на лептин е еден од начините на реализација на ефекти посредувани од страна на тирозин киназа семејството yanuskinazy (Yakovlev Цц-2). Рецептор на лептин е димер (на пример, се состои од два делови) и на лептин се врзува за екстрацелуларниот дел на рецепторот, што доведува до конформациона промени, што го прави можно да се интрацелуларната фосфорилација и активирање поврзани со Yakovlev Цц-2-молекул. Активирање Yakovlev Цц-2 молекули промовира тирозин фосфорилација на други фрагменти во рамките на лептин-рецептор-комплексот-Yakovlev Цц-2 посредува интрацелуларен сигнал.
интрацелуларен сигнали вклучуваат фосфорилација на сигнал конвертор и активатор на транскрипција - протеин кој го активира транскрипција процеси во целните гени на лептин иницирање на синтезата на протеини. Фосфорилација на Yakovlev Цц-2, исто така, доведува до активирање на други интрацелуларни ензими, како што се митоген-активиран протеин киназа и fosfatidilinozitoltrikinaza. Некои лептин влијание спроведе брзо, како резултат на тоа, активирање на интрацелуларен ензими, а другите - побавно поради потребата на синтеза на нови протеини.
Еден пример на друг механизам, широко се користат во хормонална контрола на мобилен функции е активирање на ензимот аденилат циклаза рецептор на крајот со кои се соочува во клетката следниве интеракција на надворешниот крај на хормонот на трансмембрански рецептор. Аденилат циклаза катализира формирање на цикличен АМР, предизвикувајќи различни ефекти во клетка, контролирање на својата активност. cAMP се нарекува втор гласник во ќелијата, како тој не е хормон кој посредува во дејството на хормоните на клетките.
за мал број-пептид хормони, како што е атријален натриуретичен пептид, цикличен гванозин монофосфат, не е многу различен од cAMP е вториот пренесувач во клетка, дејствувајќи како cAMP.
 Абнормалности на чорапи гени и синдром на TVH Holt-Орам. Фактори Фибробластен раст
Абнормалности на чорапи гени и синдром на TVH Holt-Орам. Фактори Фибробластен раст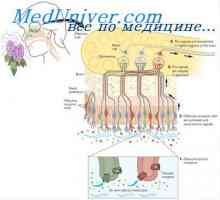 Поттикнување на мирисни клетки. Адаптација на мирисни клетки
Поттикнување на мирисни клетки. Адаптација на мирисни клетки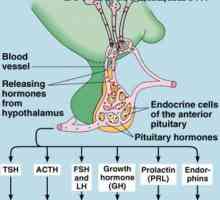 Интрацелуларна хормонски рецептори. Механизми на средните медијатори
Интрацелуларна хормонски рецептори. Механизми на средните медијатори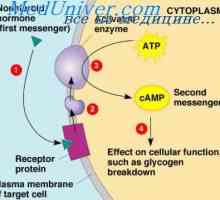 Хормонски рецептори. Бројот и чувствителноста на хормонски рецептори
Хормонски рецептори. Бројот и чувствителноста на хормонски рецептори Класи и видови на хормони. синтеза на хормони
Класи и видови на хормони. синтеза на хормони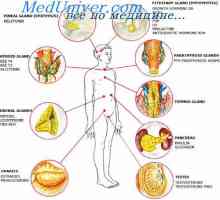 Фосфолипиди како секундарни посредници. Систем калмодулин калциум-како втор гласник
Фосфолипиди како секундарни посредници. Систем калмодулин калциум-како втор гласник На функциите на тироидната жлезда хормони. Влијанието на тироидни хормони на метаболизмот
На функциите на тироидната жлезда хормони. Влијанието на тироидни хормони на метаболизмот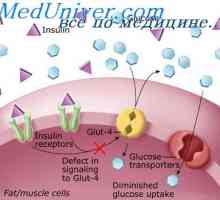 Физиологија на инсулин. Ефекти на инсулинот на мобилен
Физиологија на инсулин. Ефекти на инсулинот на мобилен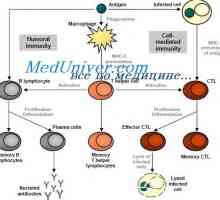 Сигнали патарина-како рецептори. Улогата на молекулата myd88
Сигнали патарина-како рецептори. Улогата на молекулата myd88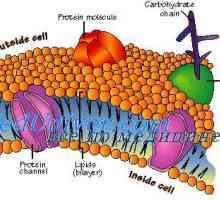 Мембрана на клетката. Во структурата на клеточната мембрана
Мембрана на клетката. Во структурата на клеточната мембрана Glycocalyx. Мобилен органели и цитоплазмата
Glycocalyx. Мобилен органели и цитоплазмата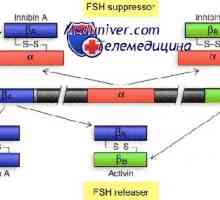 Механизмот на дејство на активин и инхибин
Механизмот на дејство на активин и инхибин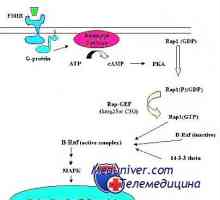 Механизмот на дејство на гонадотропин рецептори. трошење на рецептори
Механизмот на дејство на гонадотропин рецептори. трошење на рецептори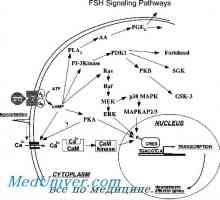 Гонадотропин рецептори. Структура и функција
Гонадотропин рецептори. Структура и функција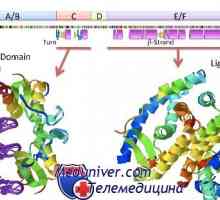 Нуклеарна рецептори за стероидни хормони: естроген, прогестерон, андроген
Нуклеарна рецептори за стероидни хормони: естроген, прогестерон, андроген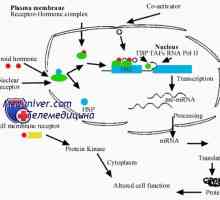 Стероид хормон интеракција со клетката. Биолошката активност на хормон
Стероид хормон интеракција со клетката. Биолошката активност на хормон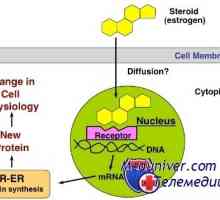 Механизмите на дејство на хормоните. рецептори
Механизмите на дејство на хормоните. рецептори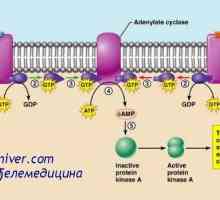 Не-геномната ефекти на стероиди. G-протеин-спарен рецептори
Не-геномната ефекти на стероиди. G-протеин-спарен рецептори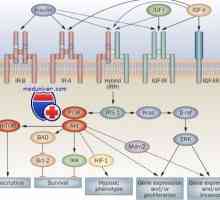 Рецептори со тирозин киназа активноста. Рецепторите за инсулин и фактори за раст
Рецептори со тирозин киназа активноста. Рецепторите за инсулин и фактори за раст Рецепторите за гонадотропна ослободувачки хормон. Агонисти и антагонисти на
Рецепторите за гонадотропна ослободувачки хормон. Агонисти и антагонисти на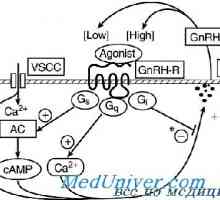 Рецептор на гонадотропин-ослободувачки хормон. Мутации GnRH рецептор
Рецептор на гонадотропин-ослободувачки хормон. Мутации GnRH рецептор
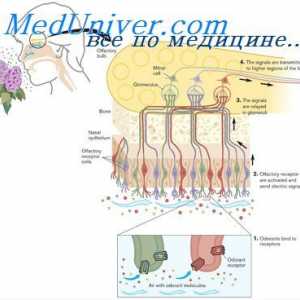 Поттикнување на мирисни клетки. Адаптација на мирисни клетки
Поттикнување на мирисни клетки. Адаптација на мирисни клетки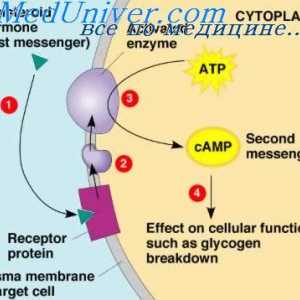 Хормонски рецептори. Бројот и чувствителноста на хормонски рецептори
Хормонски рецептори. Бројот и чувствителноста на хормонски рецептори Фосфолипиди како секундарни посредници. Систем калмодулин калциум-како втор гласник
Фосфолипиди како секундарни посредници. Систем калмодулин калциум-како втор гласник Класи и видови на хормони. синтеза на хормони
Класи и видови на хормони. синтеза на хормони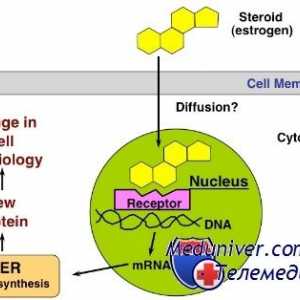 Механизмите на дејство на хормоните. рецептори
Механизмите на дејство на хормоните. рецептори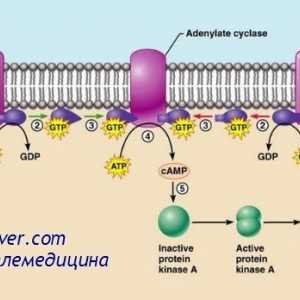 Не-геномната ефекти на стероиди. G-протеин-спарен рецептори
Не-геномната ефекти на стероиди. G-протеин-спарен рецептори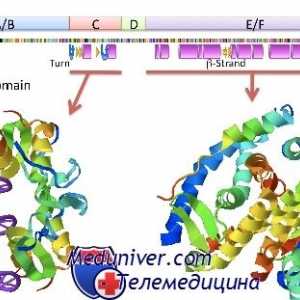 Нуклеарна рецептори за стероидни хормони: естроген, прогестерон, андроген
Нуклеарна рецептори за стероидни хормони: естроген, прогестерон, андроген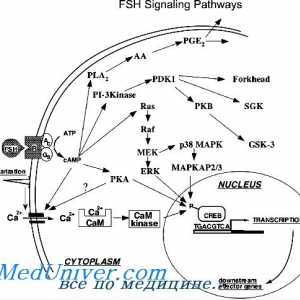 Гонадотропин рецептори. Структура и функција
Гонадотропин рецептори. Структура и функција Рецепторите за гонадотропна ослободувачки хормон. Агонисти и антагонисти на
Рецепторите за гонадотропна ослободувачки хормон. Агонисти и антагонисти на Абнормалности на чорапи гени и синдром на TVH Holt-Орам. Фактори Фибробластен раст
Абнормалности на чорапи гени и синдром на TVH Holt-Орам. Фактори Фибробластен раст